- CHIMIOSYNTHÈSES
- CHIMIOSYNTHÈSESLa chimiosynthèse est l’élaboration, par les êtres vivants, des constituants de leur protoplasme. Ils utilisent à cette fin des matériaux divers et se servent, pour en combiner les éléments, de l’énergie chimique. La chimiosynthèse s’oppose ainsi à la photosynthèse réalisée par les végétaux et par quelques bactéries à chlorophylles qui utilisent l’énergie de la lumière.Prise dans son sens le plus large, la chimiosynthèse est une des propriétés communes à tous les organismes. Les animaux n’ont d’autre source d’énergie pour réaliser leurs synthèses que celle qu’ils peuvent tirer de l’oxydation de leurs aliments. Il en est de même pour les végétaux dépourvus de chlorophylle et pour les organes non chlorophylliens des plantes. Quant aux organes chlorophylliens eux-mêmes, ils peuvent aussi, en remaniant les composés qu’ils ont synthétisés initialement grâce à l’énergie lumineuse, réaliser des chimiosynthèses.Mais la chimiosynthèse a aussi un sens plus restreint, initialement défini à la suite des travaux de Winogradsky, à la fin du siècle dernier, sur le mode de vie des bactéries autotrophes. Winogradsky a découvert que certaines bactéries sont capables d’assimiler le carbone du dioxyde de carbone tout en oxydant des composés minéraux: sulfures, sels d’ammonium par exemple. Il s’agit de bactéries incolores, les unes abondantes dans les eaux sulfureuses (sulfobactéries), les autres dans les sols (bactéries de la nitrification).Ainsi, d’après la source d’énergie utilisée par les organismes et d’après l’origine du carbone qu’ils peuvent assimiler, on oppose les phototrophes (végétaux et bactéries pourvus de chlorophylles) aux chimiotrophes ; et, parmi ceux-ci, les chimiolithotrophes qui oxydent des substances minérales et utilisent le bioxyde de carbone comme aliment carboné (bactéries autotrophes, non pigmentées) aux chimio-organotrophes , qui requièrent une alimentation carbonée nécessairement organique (animaux, végétaux non chlorophylliens, bactéries hétérotrophes). Se rapprochent de ces derniers les organismes parasites intracellulaires et les virus qui tirent énergie et aliments de leur cellule hôte.Comme les synthèses effectuées par les animaux et les autres organismes hétérotrophes sont exposées à propos de leur métabolisme [cf. MÉTABOLISME], seules seront envisagées ici les chimiosynthèses des bactéries chimiolithotrophes, qui fournissent un bon exemple du couplage d’une réaction de synthèse, l’assimilation du gaz carbonique, avec une oxydation génératrice de l’énergie nécessaire.Elles jouent un rôle important dans les cycles de l’azote et du soufre au sein de la biosphère.1. Les bactéries chimiolithotrophesSi elles assimilent toutes le carbone du bioxyde de carbone ou des carbonates, les bactéries chimiolithotrophes se distinguent les unes des autres par la nature des réactions d’oxydation énergétiquement couplées à la réduction de C2.Parmi les premières connues se trouvent les bactéries de la nitrification (Winogradsky, 1890). Ce sont des autotrophes complets, leur culture ne requiert aucun apport de substance organique, aucun facteur de croissance. Les Nitrosomonas qui oxydent les sels d’ammonium en nitrites et les Nitrobacter qui oxydent les nitrites en nitrates sont les agents de la nitrification dans les sols.L’oxydation réalisée par les Nitrosomonas (cf. tableau) est beaucoup plus exergonique, c’est-à-dire qu’elle libère plus d’énergie, que l’oxydation réalisée par les Nitrobacter .Les bactéries sulfuraires non chlorophylliennes, telles les Beggiatoa , qui sont des organismes filamenteux des eaux sulfureuses, étudiées par Winogradsky quelques années auparavant (1887), oxydent les sulfures en soufre et ce dernier en sulfates. D’autres bactéries, non filamenteuses, les Thiobacillus , oxydent soit les sulfures, soit le soufre en suspension colloïdale dans l’eau, soit les thiosulfates, et bien d’autres composés minéraux du soufre, selon les espèces biologiques.Ce sont généralement des autotrophes stricts. Le cas de Thiobacillus denitrificans est particulier: il peut vivre en présence d’air et alors se comporter comme les autres Thiobacillus ; mais, en anaérobiose, il oxyde les composés du soufre à l’aide des nitrates dont il libère l’azote à l’état gazeux.D’autres bactéries du sol oxydent les sels ferreux en sels ferriques et utilisent l’énergie libérée, pour leurs synthèses.Les bactéries qui oxydent l’hydrogène se trouvent dans les eaux riches en matières organiques en décomposition. Chez les Hydrogenomonas , l’oxydation de l’hydrogène produit de l’eau: H2 + 1/2 2H2O; très exergonique, cette réaction, explosive in vitro , est catalysée par une enzyme, une hydrogénase.Un autre type de chimiosynthèse couplée à l’oxydation de l’hydrogène est réalisé par une bactérie anaérobie stricte: Desulfovibrio desulfuricans . Ce sont alors les sulfates qui remplacent l’oxygène.2. L’assimilation du carboneLe mécanisme de la fixation et de la réduction du bioxyde de carbone par les bactéries chimiolithotrophes est très voisin du mécanisme réalisé par les végétaux à photosynthèse. Il a été étudié avec Thiobacillus denitrificans et un Hydrogenomonas , à l’aide de dioxyde de carbone enrichi en atomes radioactifs, 14C2. La radioactivité du carbone permet de suivre les différentes étapes de son assimilation en étudiant la succession des composés qui apparaissent radioactifs au cours de durées progressives.Ces étapes, catalysées par des enzymes selon le schéma ci-contre (fig. 1) se présentent ainsi: un glucide à cinq atomes de carbone, le ribulose-bisphosphate fixe C2 et se rompt en deux molécules de phosphoglycérate, corps à trois atomes de carbone. Ensuite intervient la réaction énergétiquement la plus importante: la réduction du phosphoglycérate en un glucide (triose). Cette réaction nécessite l’intervention d’un réducteur, transporteur d’électrons ou d’hydrogène, au rôle très général dans le métabolisme de tous les organismes vivants: le nicotinamide dinucléotide (NADH2 sous la forme réduite, NAD sous la forme oxydée) ou son phosphate (NADPH2). La réaction de réduction nécessite également l’intervention d’un transporteur d’énergie universel dans le monde vivant: l’adénosine triphosphate (ATP) qui la rend énergétiquement possible.Les molécules de triose formé ainsi peuvent suivre deux voies comparables à celles qui sont réalisées dans les végétaux chlorophylliens [cf. PHOTOSYNTHÈSE]. Par une série de polymérisations et d’isomérisations, elles régénèrent le ribulose-bisphosphate, fixateur initial de C2, assurant la poursuite de l’incorporation du carbone minéral dans des molécules organiques, au cours d’un véritable cycle réactionnel (cycle de Calvin, biochimiste contemporain, prix Nobel 1961).L’excédent de glucides formés suit une autre voie aboutissant à l’accumulation de réserves par les cellules ou à la synthèse d’autres constituants protoplasmiques, après de nouvelles réductions en chaînes carbonées lipidiques ou protéiques.L’entretien du pouvoir de synthèse demande que les transporteurs d’électrons NAD ou NADP soient à nouveau rechargés en électrons + protons (ou hydrogène) et que l’ATP soit également régénéré pour pouvoir intervenir à nouveau.Cette dernière régénération se fait par une réaction de phosphorylation: adénosine diphosphate + phosphate inorganique (minéral)ATP.Réductions des transporteurs et phosphorylation, réactions endergoniques, exigeantes en énergie, sont couplées aux oxydations exergoniques, libératrices d’énergie, affectant les substances minérales citées dans le paragraphe précédent.3. Les oxydations des substances minéralesLe couplage des réactions d’oxydation et des chimiosynthèses nécessite en fait le jumelage de ces oxydations avec la recharge des transporteurs d’électrons ou d’énergie.L’exemple le plus simple de la réduction du NAD en NADH2 est donné par les Hydrogenomonas , l’hydrogène moléculaire pouvant réduire le NAD. La réaction est catalysée par une enzyme, l’hydrogénase, présente dans les cellules de ces bactéries; elle est énergétiquement spontanée. Pour les mêmes bactéries, il est vraisemblable que la recharge de l’ATP est jumelée à l’oxydation d’une fraction du NADH2 régénéré.En ce qui concerne les composés soufrés, le mécanisme d’oxydation le mieux connu est celui du sulfite en sulfate, réalisé par le Thiobacillus . Un nucléotide voisin de l’ATP intervient, l’adénosine monophosphate (AMP) qui se combine au sulfite avec oxydation de ce dernier.L’adénosine monophosphosulfate (APS) peut ensuite, au cours d’une seconde réaction enzymatique, échanger son radical sulfate contre un phosphate. Il se forme ainsi de l’adénosine diphosphate (ADP). Dans une troisième étape, deux molécules d’ADP se transforment en une d’ATP et une d’AMP. Ce dernier composé est donc régénéré et disponible pour une nouvelle combinaison avec le sulfite. Le bilan comprend la formation d’une molécule d’ATP par molécule de sulfite oxydé.Des mécanismes d’oxydation de l’ion ammonium en ion nitrite, par les Nitrosomonas , et de l’oxydation des nitrites par les Nitrobacter ont été également proposés [cf. AZOTE]. Tous présentent trop de difficultés énergétiques pour que l’on puisse actuellement comprendre comment ces bactéries parviennent à obtenir le NADPH2 et l’ATP nécessaires à leurs synthèses, alors que cette nécessité même est prouvée expérimentalement.Enfin, différents transporteurs d’électrons, appartenant au groupe des cytochromes, protéines liées au fer, existent chez toutes les bactéries précédemment citées. Ils interviennent vraisemblablement lors de l’oxydation de diverses substances minérales, dans des réactions couplées à la formation de l’ATP (fig. 2).4. Le rendement énergétique des chimiosynthèsesLe rendement énergétique le plus fréquemment déterminé concerne l’ensemble des synthèses réalisées par un organisme en croissance et non un processus de synthèse particulier.Il peut être calculé de deux manières, selon que l’on considère les chaleurs de combustion (face=F0019 蓮H) des constituants formés et des composés oxydés, ou les variations des enthalpies libres (face=F0019 蓮G0) des réactions mises en jeu.Dans le premier mode de calcul, il sera défini par le rapport:
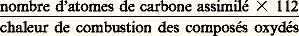 (les unités sont exprimées en kcal; 112 est la chaleur de combustion d’un atome-gramme de carbone réduit au niveau de réduction des glucides, valeur voisine de la chaleur de combustion, par atome-gramme de carbone, des organismes considérés).Le second mode, établi d’après un même type de rapport, conduit à des valeurs un peu différentes, puisque le 蓮G0 correspondant à la réduction d’une molécule-gramme de C2 est de 116 kcal, correspondant à 485 kJ et que le 蓮G0 de l’oxydation des composés oxydés diffère également de leur chaleur de combustion, la variation d’entropie qui accompagne les réactions n’étant pas nulle.Les premiers calculs faits par Winogradsky, pour la croissance des Nitrosomonas et des Nitrobacter , d’après les mesures de chaleur de combustion, donnent des valeurs de 5 à 6 p. 100. Le deuxième mode de calcul indique des valeurs de 6 à 8 p. 100. Plus récemment, divers auteurs ont trouvé des rendements de 25 p. 100 et plus, pour des cultures très jeunes de Nitrosomonas et de Thiobacillus . Ce sont des optimums qui ne se maintiennent pas lorsque les cultures vieillissent.Des valeurs comparables se retrouvent pour les bactéries hétérotrophes, avec les mêmes variations selon l’âge des cultures. Les calculs faits d’après les besoins énergétiques théoriques de la synthèse des seuls glucides conduisent à des valeurs plus élevées. Mais un organisme, même immobile en apparence, dépense pour l’absorption de ses aliments, ses excrétions, ses mouvements protoplasmiques, des quantités d’énergie difficiles à estimer qui entraînent une importante augmentation de sa consommation énergétique.5. Importance des bactéries chimiolithotrophes dans la biosphèreCes bactéries jouent un rôle fondamental, dans les sols et dans les eaux douces ou marines, à propos du recyclage de l’azote et du soufre. Ainsi, pour l’azote, leur importance est d’autant plus grande que cet élément y est relativement rare sous une forme assimilable. Les végétaux, source initiale de nourriture pour tout le monde animal, assimilent au mieux l’azote des nitrates qui, dans les sols, se trouve présent en quantités moyennes de 10 milligrammes par kilogramme de terre sèche. L’azote ammoniacal assimilable y est environ deux fois moins abondant, alors que l’azote organique est plus de cent fois plus répandu. Ainsi, les formes minérales sont-elles souvent quantitativement limitantes pour le développement des organismes.Après la dégradation des protéines des cadavres, la libération d’ammoniaque ou de sels d’ammonium fournit aux bactéries nitreuses le substrat oxydable nécessaire à leur chimiosynthèse et elles-mêmes procurent les nitrites aux bactéries nitrifiantes. Ces deux étapes de transformations minérales des substances azotées mettent les nitrates à la disposition des végétaux qui les réduisent et en intègrent l’azote dans leurs protéines. La nitrosation et la nitratation demandent l’intervention de l’oxygène, aussi l’aération des sols et des eaux est-elle importante. En conditions anaérobies, le développement de bactéries dénitrifiantes, utilisant les nitrates comme oxydants, à la place de l’oxygène, provoque des pertes d’azote heureusement compensées par les organismes fixateurs d’azote moléculaire [cf. AZOTE]. Les Thiobacillus , les sulfobactéries des sols, celles des eaux douces, jouent un rôle parallèle à celui des bactéries de la nitrification dans le cycle du soufre.Pour certaines d’entre elles, il arrive que des conditions locales particulières favorisent leur abondance. Les eaux de drainage de mines, riches en sulfures métalliques, les eaux thermales sulfureuses peuvent être très riches en sulfobactéries qui tolèrent bien l’acidité et dont certaines espèces ont une remarquable résistance aux températures de 70-75 0C.Les hydrogénobactéries sont plus rares que les précédentes en raison même de la faible fréquence de l’hydrogène libre.Toutes les bactéries chimiolithotrophes ont joué vraisemblablement un rôle géochimique important dans l’apparition sur la terre de composés oxydés, nitrates et sulfates, au cours de la vie primitive qui s’est développée originellement dans des milieux certainement beaucoup plus réducteurs que les milieux de vie actuellement les plus fréquents.
(les unités sont exprimées en kcal; 112 est la chaleur de combustion d’un atome-gramme de carbone réduit au niveau de réduction des glucides, valeur voisine de la chaleur de combustion, par atome-gramme de carbone, des organismes considérés).Le second mode, établi d’après un même type de rapport, conduit à des valeurs un peu différentes, puisque le 蓮G0 correspondant à la réduction d’une molécule-gramme de C2 est de 116 kcal, correspondant à 485 kJ et que le 蓮G0 de l’oxydation des composés oxydés diffère également de leur chaleur de combustion, la variation d’entropie qui accompagne les réactions n’étant pas nulle.Les premiers calculs faits par Winogradsky, pour la croissance des Nitrosomonas et des Nitrobacter , d’après les mesures de chaleur de combustion, donnent des valeurs de 5 à 6 p. 100. Le deuxième mode de calcul indique des valeurs de 6 à 8 p. 100. Plus récemment, divers auteurs ont trouvé des rendements de 25 p. 100 et plus, pour des cultures très jeunes de Nitrosomonas et de Thiobacillus . Ce sont des optimums qui ne se maintiennent pas lorsque les cultures vieillissent.Des valeurs comparables se retrouvent pour les bactéries hétérotrophes, avec les mêmes variations selon l’âge des cultures. Les calculs faits d’après les besoins énergétiques théoriques de la synthèse des seuls glucides conduisent à des valeurs plus élevées. Mais un organisme, même immobile en apparence, dépense pour l’absorption de ses aliments, ses excrétions, ses mouvements protoplasmiques, des quantités d’énergie difficiles à estimer qui entraînent une importante augmentation de sa consommation énergétique.5. Importance des bactéries chimiolithotrophes dans la biosphèreCes bactéries jouent un rôle fondamental, dans les sols et dans les eaux douces ou marines, à propos du recyclage de l’azote et du soufre. Ainsi, pour l’azote, leur importance est d’autant plus grande que cet élément y est relativement rare sous une forme assimilable. Les végétaux, source initiale de nourriture pour tout le monde animal, assimilent au mieux l’azote des nitrates qui, dans les sols, se trouve présent en quantités moyennes de 10 milligrammes par kilogramme de terre sèche. L’azote ammoniacal assimilable y est environ deux fois moins abondant, alors que l’azote organique est plus de cent fois plus répandu. Ainsi, les formes minérales sont-elles souvent quantitativement limitantes pour le développement des organismes.Après la dégradation des protéines des cadavres, la libération d’ammoniaque ou de sels d’ammonium fournit aux bactéries nitreuses le substrat oxydable nécessaire à leur chimiosynthèse et elles-mêmes procurent les nitrites aux bactéries nitrifiantes. Ces deux étapes de transformations minérales des substances azotées mettent les nitrates à la disposition des végétaux qui les réduisent et en intègrent l’azote dans leurs protéines. La nitrosation et la nitratation demandent l’intervention de l’oxygène, aussi l’aération des sols et des eaux est-elle importante. En conditions anaérobies, le développement de bactéries dénitrifiantes, utilisant les nitrates comme oxydants, à la place de l’oxygène, provoque des pertes d’azote heureusement compensées par les organismes fixateurs d’azote moléculaire [cf. AZOTE]. Les Thiobacillus , les sulfobactéries des sols, celles des eaux douces, jouent un rôle parallèle à celui des bactéries de la nitrification dans le cycle du soufre.Pour certaines d’entre elles, il arrive que des conditions locales particulières favorisent leur abondance. Les eaux de drainage de mines, riches en sulfures métalliques, les eaux thermales sulfureuses peuvent être très riches en sulfobactéries qui tolèrent bien l’acidité et dont certaines espèces ont une remarquable résistance aux températures de 70-75 0C.Les hydrogénobactéries sont plus rares que les précédentes en raison même de la faible fréquence de l’hydrogène libre.Toutes les bactéries chimiolithotrophes ont joué vraisemblablement un rôle géochimique important dans l’apparition sur la terre de composés oxydés, nitrates et sulfates, au cours de la vie primitive qui s’est développée originellement dans des milieux certainement beaucoup plus réducteurs que les milieux de vie actuellement les plus fréquents.
Encyclopédie Universelle. 2012.